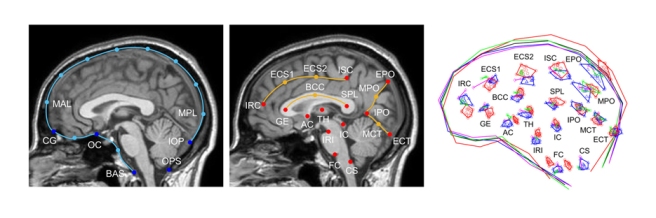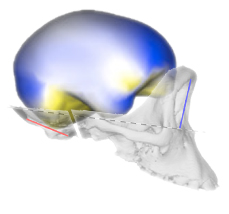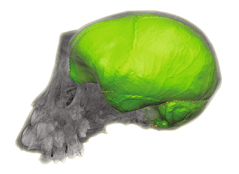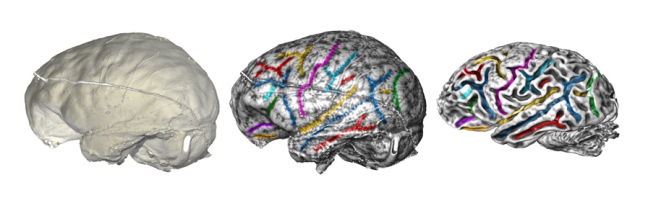
A large team led by Angela Friederici has now published a methodological article to describe a pipeline to investigate brain evolution, integrating anatomical and behavioral information from living and extinct species. To analyze correlations between brain structure and function, post-mortem brain imaging and histology in non-human primates are matched with (pre-mortem) behavioral data. Such correlations can be extrapolated to fossil endocasts. In living primates, shape analysis is employed to localize endocranial surface features, then matched with the sulcal pattern of the corresponding brain. Adult skulls are micro-CT scanned with a resolution of 90 μm. Results are, indeed, promising and exciting, revealing a very good correspondence between endocranial imprints and cortical elements. According to the authors, “almost all brain sulci leave unambiguous impressions in the endocranial cavity”. Major differences between endocranial and brain form are due to brain shrinking (mostly in the vertical dimension) after extraction, and removal of the connective tissues and cerebrospinal fluid. I guess the absence of blood pressure is another crucial factor. In the paper, language is finally used as an example to discuss how this pipeline can be employed to investigate cognitive functions that are key features in human evolution.
The large number of people, institutions, and methods necessary to handle this pipeline reminds us that approaches like this one are, indeed, very difficult to organize. Technical outputs, methodological issues, and their final interpretations represent an intricate network of variables and parameters, which require much caution when coming to the final inferences and conclusions. Nonetheless, the publication of this article also suggests that such comprehensive and efficient approaches are, at present, already available. And this is definitely the very good news.
This article represents a very useful and efficient tutorial on current comparative neuroanatomy, in particular when dealing with traditional aspects associated with evolutionary anthropology. We must anyway remark (once more) two relevant points. First, in this field, comparative approaches with humans and apes are essential. However, we should not forget that living apes are derived taxa, not ancestral ones. Furthermore, they are specialized for a kind of environment profoundly different from the one in which humans evolved. Their striking differences in cranial architecture add one more crucial problem, when they are used as models in human paleoneurology and, in general, for human brain or cognitive evolution. The second point deals with the fact that we keep focusing on human evolution as the main target of research. This is undeniably a priority, no doubt. But it should not be the only target. Apes biology and evolution are also amazing topics, to which we should probably pay more attention. And, in this case, we should probably avoid taking humans as models when investigating their peculiar derived traits.