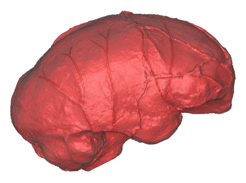
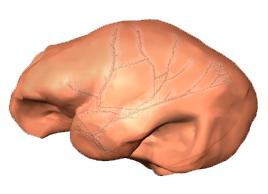
This week we have published a paleoneurological study of the specimen DAN5/P1, found in Ethiopia (Gona) and dated to 1.5 million years. According to the cranial features, the skull can be assigned to Homo ergaster/erectus. Nonetheless, the cranial capacity is very small, estimated as 598 cc. Ralph Holloway reconstructed an endocast based on the available bones. The endocranial morphology suggests brain proportions that are very similar to australopiths and to specimens generally assigned to the elusive species H. habilis, like KNM-ER 1805 and KNM-ER 1813. Most of the brain variation in these early and archaic hominids is apparently due to size, and patent differences between genera or species are difficult to establish. This can mean that most brain diversity within and between these taxa is only due to size changes and shared allometric patterns. Or that the fossil record is too scanty to provide robust statistical information. Or that endocasts, although representing the most valuable source of information on the brain of extinct species, cannot show subtle cortical details. Either way, we can probably say that macroscopic brain differences between these early and archaic species (beyond mean brain size) are absent or at least subtle. Of course, the absence of (anatomically or statistically) consistent macroscopic difference does not exclude the possibility of internal changes, associated with cell and tissue organization, connections, metabolisms, or neurotransmitters. The difficulties in detecting specific brain differences in these early humans remark the uncertainties on the phylogenetic origin of our genus, as mentioned in this recent article on human paleoneurology. Here’s another perspective review, pointing at the possibilities and limitations of the field.